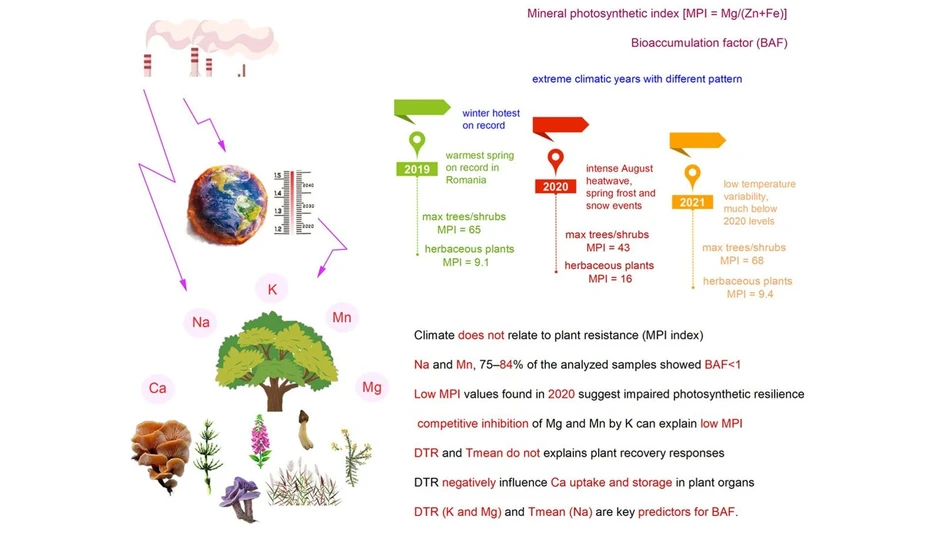
Nutrient imbalances caused by industrial pollution were not affected by extreme climate events
Journal of Hazardous Materials Advances 21, 2026, 101071: Graphical abstract
This study investigates how industrial pollution and extreme climate conditions affect nutrient cycling and plant mineral balance in the polluted region of Copșa Mică, Romania. Analysis of plants, fungi, soils, sediments, water, and litter from 2019–2021 revealed persistent nutrient imbalances, particularly involving potassium, magnesium, and manganese.
Results suggest that industrial contamination exerts a stronger influence on nutrient dynamics than short-term climate extremes. Biochemical indicators pointed to reduced plant resilience and weakened biogeochemical relationships, emphasizing the need for long-term monitoring and sustainable management of polluted ecosystems under changing environmental conditions.
The original article
Nutrient imbalances caused by industrial pollution were not affected by extreme climate events
Constantin Nechita, Andreea Maria Iordache, Carmen Roba, Tomas Pluhacek, İsmail Koç, Roxana Elena Ionete, J. Julio Camarero
Journal of Hazardous Materials Advances 21, 2026, 101071
https://doi.org/10.1016/j.hazadv.2026.101071
licensed under CC-BY 4.0
Selected sections from the article follow. Formats and hyperlinks were adapted from the original.
Industrial and urban activities are a significant source of pollution in soil, water, and living organisms, impacting ecosystems (Ali et al., 2021a; Boukich et al., 2025a; Iordache et al., 2020; Iordache et al., 2024a; Iordache et al., 2024b). Severe human and environmental issues occur due to toxicity, persistence, and non-degradability of mineral elements (Ben-tahar et al., 2025b; Kumar et al., 2020; Souza et al., 2021). Systematic pollution from multiple sources, combined with climate change, negatively affects terrestrial and aquatic resources through contamination (Boukich et al., 2025b; Boukich et al., 2025c; Chowdhary et al., 2020; Iordache et al., 2022a). However, we still do not know how pollution interacts with extreme climate events impacting the distribution of essential minerals in ecosystem components.
Essential minerals are required for the functioning of biological processes, such as activating enzyme reactions and stabilizing cation (e.g., Mn and Mg) exchange (Ben-tahar et al., 2025a; Li et al., 2021). Macro minerals (e.g., Na, Mg, K, Ca) maintain the ionic balance of structural compounds, amino acids, and nucleic acids, and are associated with toxicological effects after interactions with biological systems (Ali, 2023). They act as cofactors of vital enzymes involved in physiological and biochemical processes. The synergistic and antagonistic interactions among minerals in plants reflect cumulative effects and irreversible damage, as seen with Mn vs. K (Xie et al., 2021); however, the primary focus remains on critical nutrient concentrations (Jin et al., 2023).
Assessing changes in essential minerals is key for determining ecosystem pollution levels. For instance, the mineral photosynthetic index (MPI = [Mg/(Zn+Fe)]) is used to assess tolerance to heavy metals and abiotic stress by reflecting the balance between key photosynthetic processes, particularly chlorophyll activity (Mg) and electron transport (Fe, Zn) (Diatta et al., 2021). Deficiencies in these nutrients reduce electron flow and the maximum quantum yield of photosystem II. Mg and Fe are essential for photosynthesis, while Zn and Fe support antioxidant defense and nutrient stability (Shinozaki and Yoshimoto, 2021; Singh et al., 2024). Imbalances suggest higher stress vulnerability or tolerance (Rahim et al., 2025). Mg acts directly in photosynthesis by supporting chlorophyll structure and activating enzymes such as Rubisco, and indirectly supports protein synthesis and reactive oxygen species scavenging (Jeevanraj et al., 2025). A high Zn/Fe ratio indicates effective downstream processes and a strong antioxidant system, showing resilience to stress (Ali et al., 2021b). Overall, this ratio indicates how metabolic functions are affected by mineral nutrition on photosynthesis and stress response (Saudy et al., 2022).
However, there remains a lack of global monitoring studies to understand the behavior and evolution of macronutrients in plants following the closure of industrial facilities and under current warming trends, which can increase vegetation stress. Earlier research focused on toxic metals (Hg, Cd, and As), overlooking the roles of nutrients (Bartha et al., 2020; Harja et al., 2023; Hoaghia et al., 2022; Nechita et al., 2021; Vlad et al., 2019). K, Ca, Mg, Mn, and Na can function as mediators in heavy metal toxicity and act as indicators of biogeochemical remediation, which is essential for nutrient management to improve plant resistance to environmental stress. Most studies used retrospective approaches based on tree rings to evaluate changes in growth resilience (Nechita et al., 2021; Nechita et al., 2025a). An alternative methodology for evaluating plant recovery after environmental stress employs the MPI, which enables assessment across multiple substrates (trees, shrubs, and herbaceous plants) (Diatta et al., 2021). In this context, the objectives of this study were to: (i) evaluate the contents of macro minerals (K, Ca, Mg, Mn, and Na) in 40 environmental substrates or matrices (plants, fungi, water, soil, and sediments), (ii) find patterns between macro elemental content and climatic variables over three years of monitoring (2019‒2021), (iii) evaluate the enrichment and bioconcentration factors in soil and plants, and (iv) model the mineral photosynthetic index as a function of climatic factors to assess recovery patterns. This study aims to assess the effects of mineral deficiency and imbalance resulting from climatic stress on plant mineral nutrition and resilience in a highly industrialized and polluted region.
2. Materials and methods
2.2. Sample preparation and analysis
The content of essential minerals was determined using the recently published inductively coupled plasma optical emission spectroscopy (ICP-OES) method (Iordache et al., 2022b; Iordache et al., 2024a; Iordache et al., 2024b; Nechita et al., 2025b). Briefly, biotic and abiotic samples were digested in a closed-vessel microwave digestion unit (MARS6, CEM Corporation, Matthews, NC, USA, One Touch) using a mixture of concentrated acids according to either the US EPA 3015, the NPDES, or sand digestion methods. Total macro mineral levels were measured using an ICP-OES Plasma Quant 9100 Elite (Analytik Jena, Germany) with five-point external calibration. All ICP-OES measurements were performed in five technical replicates. The ICP-OES method was validated for linearity, limit of detection, trueness, and precision. The Standard Reference Material (SRM) 2710a Montana I soil, NIST 1570A (trace elements in spinach), and BRC679 (white cabbage trace elements) were used for the ICP-OES method validation in terms of trueness and precision. Moreover, they served as QC samples, which were regularly analyzed within each batch to ensure the reliability of the acquired data. Detailed information on the ICP-OES parameters, method validation, and quality control samples is provided in the Supplementary Material.
3. Results
3.5. Climate conditions and changes in minerals
The content of macro minerals in environmental matrices changed over time in relation to climatic variables (Fig. 4a–c). The cumulative inertia on the first component was high and increased in 2020 and 2021. Additionally, in 2019, macro minerals showed a positive correlation with Tmean and R, and a negative correlation with DTR. In 2020, R and Ca, Mg, and Na formed a group with a positive relationship among them and were negatively associated with maximum temperature. Only Ca, Na, and K were negatively influenced by Tmax in 2021.
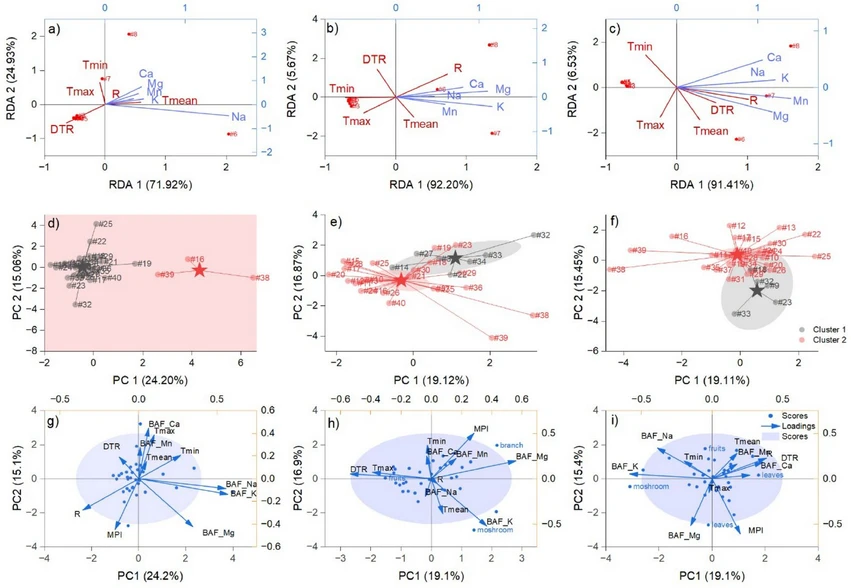 Journal of Hazardous Materials Advances 21, 2026, 101071: Fig. 4. The graphical representation of RDA analysis to assess the macro minerals relationship with climatic variables in the period 2019‒2021 (a‒c). The labels annotated with # followed by a number represent the forty matrices evaluated, as described in the Materials and Methods chapter.
Journal of Hazardous Materials Advances 21, 2026, 101071: Fig. 4. The graphical representation of RDA analysis to assess the macro minerals relationship with climatic variables in the period 2019‒2021 (a‒c). The labels annotated with # followed by a number represent the forty matrices evaluated, as described in the Materials and Methods chapter.
Segmentation of the biota dataset and dimensionality reduction revealed a low degree of climate inference in macro-mineral bioaccumulation and photosynthetic resilience (Fig. 4d–f). ANOVA results indicated that variables with significant contributions to the formation of K-means clusters were the MPI (F = 120, 68 and 66 in 2019, 2020, and 2021, respectively; p < 0.0001 in all cases), Tmean (F = 13, p < 0.05 in 2019), BAF Mg (F = 14 and 4.5 in 2020 and 2021, p < 0.05 in both cases), and Tmin (F = 4.1, p < 0.05 in 2021). The PCA analysis indicated that MPI and precipitation had negative loadings in 2019. In 2020, MPI exhibited positive loadings, whereas climatic variables did not consistently load positively or negatively on PC1 and PC2. Different loading was observed in 2021 for MPI, with PC1 being positive and PC2 negative.
4. Discussion
4.3. Nutrient imbalance and photosynthetic impairment
Climate change impacts on vegetation can interact with nutrient and water availability (Kumar et al., 2021; Palacio et al., 2018). The low MPI values found in 2020 suggest impaired photosynthetic resilience, mainly due to chronic nutrient deficiencies resulting from soil degradation. Only in the abiotic samples the DTR was associated with Ca. Our models indicate that climate is an additional stressor, but only for bioaccumulation, although its influence varied from year to year. The BAF had low values in tree and shrub organs, indicating limited bioaccumulation. Higher BAF values observed in 2021 were exceptional and were only found in Q. pubescens primary branches for K and Na. For Na and Mn, 75‒84% of the analyzed samples showed BAF<1, indicating low bioaccumulation. However, organ characteristics also contributed to the observed variability in BAF. This pattern was also noted for MPI, with estimated marginal means higher in leaves than in other organs, attributable to photosynthetic capacity. The observed elemental content patterns are consistent with the hypothesis that the competitive inhibition of Mg and Mn by K is associated with a significant decline in the MPI index in 2020 and 2021. Similar results have been reported in the literature, in which 77% of deciduous species exhibited low to moderate photosynthetic recovery, attributed to acid rain leakage (Diatta et al., 2021). Trees experience various functional impairments when they lack essential elements, such as oxidative stress, phloem cell blockage, meristem damage, and inactivation of stress-protection genes (Fromm, 2010; Nechita et al., 2025c). Furthermore, the mobility of chemical elements in plants varies significantly with age and across species (Setälä et al., 2017). Additionally, the Mg-charged particulate matter, resulting from high traffic intensity, accumulated in large amounts on the northern side (Erdem et al., 2024).
Although the GLMs indicate that climatic variables have weak explanatory power for the MPI, they significantly affected the bioaccumulation factor of K, Mg, and Na between organs. This study shows that extreme climate events do not primarily drive plant mineral photosynthetic recovery responses, but can influence elemental uptake and translocation between plant organs. Still, the low R² values of GLMs suggest that other unmeasured factors may be relevant, even though we found statistically significant effects on the bioaccumulation factor, as with DTR for K and Mg, and Tmean for Na. These unmeasured factors likely include finer-scale soil chemistry (e.g., pH, organic matter, cation exchange capacity), microclimatic conditions, plant genotype, and legacies of historical soil pollution.
The BAF values in herbaceous plants were close to 1, indicating a low bioaccumulation potential for Ca, Mg, and Na. Mn content was extremely low (near the limit of deficiency) in E. arvense and P. vulgaris in all three years of monitoring (47.9 ± 15.0, 27.6 ± 4.7 mg kg-1). In this case, the BAF was below 1. Conversely, another pattern emerged among mushrooms in 2021, when L. torminosus exhibited exceptionally high levels of macro minerals, a phenomenon not previously documented in the literature. This indicates its predisposition to absorb substantial amounts of K, Ca, Mg, and Mn; however, the BAF factor also suggested limited enrichment. Nevertheless, the disparity in mole ratios (K/Ca, K/Mg) was evident among herbaceous plants, indicating disequilibrium in plant mineral content attributable to pollution. Pollution and drought can cause mineral deficiencies by interfering with their uptake, thus disrupting the nutrient balance (Ahanger et al., 2016; Czaja et al., 2020). The availability of essential nutrients (N, P, and K) I, therefore, closely linked to geochemical processes that can interfere with their cycling, particularly in urban and industrialized areas (Macci et al., 2025).
Here, we present an alternative to retrospective approaches to assess changes in pollution impacts on ecosystems using, for instance, dendrochronology. The presented approach is limited to in situ monitoring and requires numerous analyses of annually collected samples. However, it offers a significant advantage by reflecting changes not only in the wood (tree rings), but also in several abiotic and biotic components of ecosystems. The results presented in the current study are consistent with those obtained by the dendrochronological approach in this area, based solely on tree-ring data from oak trees (Nechita et al., 2021; Nechita et al., 2025a). In the current approach, we documented insights into the macro-mineral implications for reduced resilience, using nutritional proxies for trees, shrubs, herbaceous plants, and mushrooms. Several limitations of the current methodology should be addressed in future studies, including the need to measure additional parameters (e.g., water and soil pH, cation exchange capacity, stable isotopes). Additionally, this methodology requires a robust interdisciplinary approach to interpreting interactions between abiotic stressors and plant nutrition.
5. Conclusions
We aimed to assess in situ ecosystem resilience under current climate change and historical pollution stressors, using mineral (K, Ca, Mg, and Na) deficiency and imbalance as indicators. Results indicated a high K content, supporting the hypothesis that an imbalance with other minerals disrupts the equilibrium of essential elements. Levels of macro minerals in plants varied by year, organ, matrices, and location. Compared to optimal ranges, essential nutrients were deficient in all matrices. The EF showed depletion or minimal enrichment in the litter and 25-cm deep soil. Except for K, plant element levels were low, and BAF values indicated limited bioaccumulation. In 2020, characterized by extreme climatic events, the MPI was low, suggesting that climate extremes may exacerbate stress induced by mineral deficiencies. Although long-term environmental pollution is characterized by a decreased mineral content, our findings indicate a gradual increase since 2019 following the cessation of industrial activities; however, MPI and BAF remain low, and the mineral ratio accentuates biochemical imbalance. Fitted models showed a weak link between BAF values of K versus Mg and Na with the diurnal temperature range and the mean temperature, respectively. Our findings highlight the need for improved monitoring of macronutrients in polluted areas under climate-warming conditions.